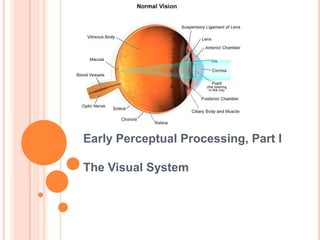
2. The Retina The vertebrate retina has ten distinct layers Inner limiting membrane Nerve fiber layer Ganglion cell layer: Layer that contains nuclei of ganglion cells and gives rise to optic nerve fibers Inner plexiform layer Inner nuclear layer Outer plexiform layer : In the macular region, this is known as the Fiber layer of Henle Outer nuclear layer External limiting membrane : Layer that separates the inner segment portions of the photoreceptors from their cell nuclei Photoreceptor layer: Rods / Cones Retinal pigment epithelium
3.
4. Neural Processing and Early Vision Each retina contains 126 million receptors 2 kinds 120 million Rods 6 million Cones
5. Receptor Cells Humans have two receptor cell types: All receptor cells have outer segments (containing the photoreactive compounds necessary for converting light energy into chemical energy), an inner segment, a perikaryal region, and a terminal Rods will have long outer segments and small, spherically shaped terminals (spherules) Cones often have a shorter outer segment, a fatter inner segment and a larger terminal (pedicles) than rods Note: In the human, foveal cones appear much more like rods than the parafoveal and peripheral cones
6. Cellular Organization All vertebrate retinas are organized according the same basic plan (i.e., two synaptic layers) Outer & Inner nuclear layers: contains photoreceptors and preprocessing neural elements Ganglion cell layer: Contains some preprocessing neural elements and the ganglion cells
7. Cellular Organization Although only the photoreceptors are the light sensitive component (all visual responses are initiated by the photoreceptors), the retina has five other basic classes of retinal neurons: Horizontal Cells Amarcrine Cells Bipolar Cells Innerplexiform Cells Ganglion Cells
8. Cellular Organization The cell bodies (perikarya) of the photoreceptors are located in the outer nuclear layer The perikarya of the Horizontal cells lie along the outer margin of the inner nuclear layer Bipolar perikarya are predominantly located in the middle layer Amacrine & Innerplexiform cell perikarya are located along the proximal border of the inner nuclear layer Ganglion cell perikarya comprise the most proximal layer, the ganglion cell layer
10. Spatial Summation 126 million photoreceptors, but only 1 million ganglion cells In some parts of the eye a single ganglion cell is receiving inputs from many photoreceptors This has implications for visual sensitivity and acuity
11. Spatial Summation The degree of convergence among photoreceptors onto ganglion cells increases with retinal eccentricity (moving from the center out into the periphery) Foveal cones have a 1:1 or near 1:1 relation with ganglion cells Receptors in the far periphery can have up to a 400:1 relation with a ganglion cell
12. Spatial Summation The concept of the ReceptiveField A region of space (visual, auditory, somatosensory) that is associated with a particular response In this case, the “region of space” is a section of retina, and the “response” is ganglion cell activity As the degree of convergence increases, so does the size of the receptive field
13. Spatial Summation Implications for visual sensitivity With a larger receptive field, more light can be caught With more photoreceptors active and pooling their collective responses onto a single ganglion cell, it is much more likely for the ganglion cell to fire With a smaller receptive field, less light can be caught With fewer photoreceptors active, it is less likely for the ganglion cell to fire
14. Spatial Summation Implications for visual acuity Large receptive fields pool light information from a large area Small details get lost It does not matter where in the receptive field light has fallen, only that enough of it has The fovea has very small receptive fields, and good acuity The peripheralretina has larger receptive fields, and poor acuity
15.
16.
17.
18.
19.
20.
21.
22.
23.
24. Receptive Fields & Inhibition Recall the concepts of excitation and inhibition Excitation = more activity Inhibition = less activity Within a receptive field, some photoreceptors are excitatory, whereas others are inhibitory to the ganglion cell
26. Center-Surround Antagonism Lateral Antagonism As you increase the size of the spot of light on the entire receptive field, the firing rate of the ganglion cell changes
27. Receptive Fields & Inhibition Implications for visual perception Mach bands The perception of light and dark bands near the borders between light and dark areas
30. What is the benefit? Although LA can lead to some interesting visual illusions, it does serve very important functions E.g. Contrast enhancement at edges Your eye is not a perfect optical instrument Edges are blurred to some degree By enhancing the difference between a light and dark region, your visual system is able to compensate for the blurring
32. The Optic Nerve Neurons leave the eye via the optic nerve Highly myelinated and therefore neural conduction is very fast Diseases that affect the myelination of neurons exhibit their symptoms here first (e.g., multiple sclerosis)
33. The Pathway to the Brain After leaving the eye… The optic nerve crosses at optic chiasm Optical fibers extend away from the optic chiasm Travel to lateral geniculate nucleus(dLGN) Optical radiation extends away from the dLGNbranch out and enter the primary visual cortex AKA striate cortex Signals are then rerouted to higher brain areas (extra-striate cortex)
34. Optic chiasm Why the crossover? Contralateral processing Right visual field is processed in the left hemisphere of the brain, and vice versa
35. The lateral geniculate nucleus The dLGNhas circular receptive fields with a center-surround configuration (more on this later) Retinotopicmap Adjacent points on the retina are adjacent points at the dLGN
36. The lateral geniculate nucleus Organized into 6 layers the eye on the same side sends information to layers 2, 3 and 5 (ipsilateral) the eye on the opposite side sends information to layers 1, 4 and 6 (contralateral) Just remember that 2 + 3 = 5 whereas 1 + 4 does not equal 6
37. The Primary Visual Cortex Contralateral processing The LEFT hemisphere processes information from the RIGHT visual field The RIGHT hemisphere processes information from the LEFT visual field Retinotopicmap Cortical (foveal) magnification Fovea is only 0.01% of retinal area, but 8-10% of the cortical area (1000x magnification)
38. The Primary Visual Cortex The visual cortex is topographically organized A large area of the visual cortex is mapped to the central portion of the visual field (retina) Retinotopic map & Cortical magnification
39. Cerebral Cortex Number of neuronal cells in cerebral cortex neurons ----------- 10-15 billion glial cells ---------- 50 billion
41. Molecular Layer External Granular Layer External Pyramidal Layer Internal Granular Layer Line of Gennariin area 17 Internal Pyramidal Layer Polymorphic Layer Golgi NisslWeigert
42. The “What” Pathway Information leaving the striate cortex and entering the temporal lobe is associated with object recognition Ventral pathway Ablating or damaging this pathway results in impairments of visual object recognition E.g. Agnosia
43. The “Where” & “How” Pathway Information leaving the striate cortex and entering the parietal lobe is associated with object location (spatial processing) Dorsal pathway Ablating or damaging this pathway results in impairments of visual object localization
44. The origins of these pathways Dorsal and ventral pathway separation begins at the level of the retina M-cells P-cells
45. The Magnocellular pathway M-cells Retinal ganglion cells with large cell bodies Innervate layers 1 and 2 of the dLGN Fed mainly by rods (rod dominated) Associated mainly with motion perception Signals in fast and transient bursts
46. The Parvocellular Pathway P-cells Retinal ganglion cells with small cell bodies Innervate layers 3, 4, 5 and 6 of the dLGN Fed mainly by cones (cone dominated) Associated mainly with color, texture and depth perception Signals in slow and sustained fashion
48. Parvo & Magno Pathways In primates, it is clear that the retinocortical visual system is organized into two major pathways (parvo & magno) The neurons which comprise these two pathways have different sensitivities to chromatic, spatial and temporal stimuli
51. Functions of the M & P Pathways Behavioral studies in monkeys Legion of the parvocellular region of the dLGN: Color vision is severely diminished High frequency spatial vision is poor Interestingly, high frequency flicker detection remains largely unaffected Legion of the magnocellular region of the dLGN: Color vision is largely retained Normal contrast sensitivity to high spatial frequencies Profound loss of sensitivity to high frequency flicker
53. The Striate Cortex Named so because of the dense plexus of geniculate axons that form distinctive bands The “primary” band is referred to as the line of Gennari (layer 4B)
55. The striate cortex Conventionally divided into 6 layers Layer 1 is near the cortical surface; layer 6 is adjacent to the white matter
56. The striate cortex The primary destination of the axons from the LGN is to layer 4, although some fibers will extend to layers 1, 3 and 6
57. The striate cortex Organization might be very similar to the retina and LGN Evidence for presence of horizontal (lateral) connections between areas of V1 Physiological evidence suggests that they are largely inhibitory
58. The Striate Cortex The striate cortex sends axon projections to the extrastriate cortex This region of the visual cortex is not distinguished by a line of Gennari Extrastriate areas include: Visual Area 2 (V2) Visual Area 4 (V4) Inferotemporal Cortex (IT) Mediotemporal Cortex (MT; V5)
59. The “Flow” of Information Recall that the magno and parvo-cellular systems have been considered largely distinct This is true, however, at higher cortical areas more and more information begins to “cross-over” Obviously, this is important for a complete sensory integration, or else we could see colors, but never associate them with a particular object, per se
60. The “Flow” of Information Not only does the striate cortex project “forward” through the visual system, but it also projects “backward” Reciprocal projects are sent back to the dLGN and superior colliculus
61. The “Flow” of Information These “backward” projections originate from the deeper layers of the cortex I.e., Layers 5 & 6 “Forward” projections tend to originate from the superficial layers
62. The “Flow” of Information Interestingly, information can then flow back to the cortex forming a feedback loop The ratio of feed-back to feed-forward projections can be as high as 900:1!
63. The Striate Cortex Very little was understood about the primary visual cortex Although it had been known that concentrically organized cells could be found at the level of the retina, these structures could not be found in V1 Stimuli optimized for a circular configuration were not “activating” cells in the visual cortex…
64. The Striate Cortex Hubel & Wiesel (1959; 1962) made a critical discovery when making extracellular recordings from single cells in the cat striate cortex Essentially, they discovered neurons that were most sensitive to elongated stimuli such as bars or edges These cells were then divided into two general categories: Simple cells Complex Cells
66. Cortical cells Simple cells Respond best to a bar of light in a particular orientation Complex cells Respond best to a bar of light of a particular orientation moving in a particular direction End-stop cells Respond best to bars of a specific length, or to joined lines (corners) moving in a specific direction
67. The striate cortex Neurons are specialized to respond best to very specific aspects of a stimulus Orientation, size, direction of movement, etc. Arranged in a side-by-side fashion, not center-surround fashion Will respond best to bars of light that share the same orientation as the receptive field
68.
69. Simple Cells It is thought that that simple cell receptive fields are formed by the addition of dLGN receptive fields that lie along a straight line This is known as hierarchical or serial processing
70. Complex Cells Like the simple cells, complex cells respond best to elongated stimuli of a particular orientation However, unlike simple cells, the object can be located anywhere within the receptive boundaries of the field
71.
72. Complex Cells Moreover, many complex cells are characterized by a sensitivity to a stimulus moving in a particular direction A stimulus will all the proper characteristics, but moving in the direction opposite to that which the cell is sensitive will illicit no response Complex cells also cannot be divided into discrete excitatory or inhibitory regions Suggests that the integration of multiple simple cells is a non-linear process
73. End-Stop Cells Originally classified as hypercomplex cells Sensitive to the length of an edge or bar stimulus It was later determined that this is a characteristic of many of the earlier cells, and so does not define a new class of cell Nevertheless, Hubel & Wiesel’s work conclusively demonstrated that hierarchical processing is central to processing of visual information As information is conveyed to higher and higher vision centers, the neurons become increasingly more specific in their response
74. Striate Cortical Architecture Many cortical neurons are binocular (receive input from both eyes) Binocular cells may mediate stereopsis The receptive fields of many binocular cells do not overlap with the sameregions in the eye Permits the coding of retinal disparity
75. Striate Cortical Architecture Many cortical neurons are binocular (receive input from both eyes) Binocular cells may mediate stereopsis The receptive fields dooverlap when an object islocated at a critical distancefrom the eyes
76. Striate Cortical Architecture Ocular dominance Columns Ocular dominance is laid out in a regular pattern of alternating right and left ocular dominance slabs A complete set of ocular dominance columns and orientation columns forms a hypercolumn(2x2mm)
77. Parvo and Magno Pathways Recall that the segregation of the parvo and magno systems persists through to V1, and somewhat beyond Staining the cortex for cytochromeoxidase reveals an irregular pattern of “blobs” within the superficial layers of the striate cortex Stripes adjoining area V2 can also be seen
78. Parvo and Magno Pathways Blobs are rich with concentrically organized, double-color opponent neurons Blobs are also connected to the stripes in V2 and this constitutes a continuation of the color sensitive parvo pathway The superficial layers between the blobs are cleverly known as the interblob region
79. Parvo and Magno Pathways The magno pathway appears to bypass the blob and interblob regions by synapsing with deeper layers of the cortex The magno pathway projects to the stripes in V2 and then to V5 (motion)
80. Cortical Modularity Evidence that specific brain regions respond best to particular classes of stimuli comes from legion and imaging studies With regard to brain imaging studies: When subjects are presented with particular kinds of visual stimulation, it is found that distinct regions of the brain become “active” and/or inhibited
81. Cortical Modularity Higher visual areas appear to have specialized functions Visual Area 4 (V4): responds to chromatic stimuli and is therefore involved in color perception Inferotemporal Cortex (IT): responds to complex forms (e.g., faces), indicating a role in form perception Both of these brain regions are considered to be a part of the parvo (“What”) stream
82. Cortical Modularity Higher visual areas appear to have specialized functions Visual Area 5 (V5): responds to motion stimuli Therefore, this brain region is associated with the magno (“Where”) stream
88. Visual Area 5 This region of the brain is heavily studied and is probably better understood than other regions Neurons in V5 respond to global movement Motion Aftereffect
90. Why is hearing important The first sense to develop The first sound you hear is your mother’s heart beat Infants react to brady-tachycardia Used in poems/music, iambic pentameter (Wagner’s 9th symphony)
91. Why is hearing important Approximately 2 million people are profoundly deaf One of every 1000 infants is born totally deaf 28 million Americans have hearing loss 80 percent of those affected have irreversible and permanent hearing damage Average age of diagnosis, 3 years of age 15 percent of the U.S. population is affected by tinnitus Presbycusis affects 1/3 of the U.S. population over 65 Estimated cost of care, 56 billion per year
102. Sound Physical definition Sound is rhythmic pressure changes in air or some other medium Perceptual definition Sound is the experience we have when we hear something
103. The Cochlea The cochlea is the organ of the inner ear responsible for transducing the auditory stimulus into electrochemical signals Small fluid filled bony structure rolled upon itself (2¾ turns)
104. The Cochlea The base is the portion of the cochlear nearest to the oval window, whereas the apex is the area furthest from the oval window Divided into two halves by the cochlear partition which extends the length of the inner cochlea Top half: scalavestibuli Bottom half: scala tympani On the cochlear partition is a structure called the organ of Corti
106. The Organ of Corti Supported by the Basilar Membrane Contains the hair cells which are the receptors for hearing Cilia protrude from the tops of the cells and are responsible for converting vibrations into electrical signals Outer hair cells Inner hair cells The tectorial membrane extends over the hair cells
107.
108. Auditory Transduction Transduction starts with the bending of the cilia What causes the bending? The in-and-out movements of the stapes on the oval window generates pressure changes in the fluid within the cochlea The rhythmic pressure changes cause the cochlear partition to vibrate up-and-down
109. Auditory Transduction Up-and-down motion of the partition Causes the Organ of Corti to vibrate Causes the tectorial membrane to move back-and-forth These two motions cause the cilia of the inner hair cells to bend because of their movement against the surrounding liquid and because they are in contact with the tectorial membrane
110.
111. Auditory Transduction The necessary amount of bending of the cilia to generate an electrical signal is VERY small 100 picometers To put this in scale, if you increased the size of a cilium to be the same as the Eiffel Tower, the minimal displacement would be 10mm
117. The Cochlea IHCs send signals to fibers which bundle together and become the auditory nerve 95% of the auditory nerve caries signals from the IHCs OHCs also send signals through the auditory nerve 5% of the auditory nerve caries signals from the OHCs
118. Békésy’s Place Theory Tonotopic Map An orderly map of frequencies along the length of the cochlea Placing electrodes at different places along the length of the cochlea and measuring the electrical response to different frequencies of sound indicates that some areas respond best to low freqs, and other areas to high freqs Specifically, the base of the cochlea responds best to high freqs, and the apex responds best to low freqs
119. The Motile Response Although the inner hair cells are responsible for transducing the auditory stimulus into electrical signals, the outer hair cells also play an important role Movement of the outer hair cells affects the movement of the basilar membrane Different outer hair cells will respond to different frequencies (high freq = base, etc.) Movement of the outer hair cells amplifies the motion of the basilar membrane and sharpens its response to specific frequencies Ultimately, this helps the ear distinguish between two very similar tones (ex: 400 vs. 405Hz)
120.
121. Auditory Pathway To the Medial geniculate nucleus (MGN) in the thalamus From the Thalamus (MGN) signals are send to the primary auditory cortex (A1) in the temporal lobe SONIC MG Superior OlivaryNucleus; Inferior Colliculus; Medial GeniculateNucleus
122.
123. The Primary Auditory Cortex (A1) Tonotopic organization Frequency analysis (perceiving pitch) Binaural integration Auditory localization
125. From A1… The auditory system (like the visual system) has What and Where pathways What stream Auditory ventral pathway Passes to the pre-frontal cortex Associated with identifying sounds Where stream Auditory dorsal pathway Passes to the parietal cortex Associated with identifying the location of sounds
126. Auditory Localization Coordinate system Azimuth – horizontal (left to right / side to side) Elevation – vertical (top to bottom / up-down) Distance – how far away is the sound
127. Auditory Localization People with normal hearing tend to be fairly good at identifying the location of sounds Listeners can localize sounds from directly in front of them most accurately (within 2 - 3.5 deg) Listeners are least accurate localizing sounds that are off to the side or behind their head (error = 20+ deg)
128. Auditory Localization Binaural Cue Auditory cue based on sounds reaching both ears Monaural Cue Auditory cue based on sound only reaching one ear
129. Binaural Cues Based on a comparison of sound signals reaching the left and right ears Sounds to the side of a listener will… …reach one ear sooner than the other Interaural time differences (ITD) …be more intense in one ear than the other Interaural intensity differences (ILD) Primarily involved in azimuth judgments
130. Down to 10 microseconds Interaural Time Difference
133. Computation of Spatial Localization The brainstem computes spatial location of sounds by using Delay Lines and cells called Coincidence Detectors These structures are able to detect the difference in arrival time of a sound to each ear Certain cells in the brainstem respond preferentially to particular timing discrepancies between the two ears
134. Computation of Spatial Localization Medial SuperiorOlivary Nucleus(MSO) Left Cochlear Nucleus A B C Right Cochlear Nucleus Delay Line Cells A, B & C are Coincidence Detectors: Activated most strongly when signals from BOTH cochlear nuclei arrive at the same time.
135. Acoustical Illusions The McGurk Effect Our perception of speech is multimodal, that is, it can be affected by many different senses The McGurk Effect illustrates how visual information can affect auditory perception Ss hear / ba-ba /, but see a person saying / ga-ga / The resulting perception is / da-da/ McGurk Link